Search for articles:
Yuqiao Dong , Soumya El Abed
, Soumya El Abed
Corresponding authors:
Received: 2019-05-17
Revised: 2019-06-14
Accepted: 2019-06-29
Online: 2020-01-15
Copyright: 2020 Editorial board of Journal of Materials Science & Technology Copyright reserved, Editorial board of Journal of Materials Science & Technology
About authors:
1 These authors contributed equally to this work.
More
Abstract
Algae are reported to be corrosive, while little is known about the role of the algae associated bacteria in the corrosion process. In the present study, Halomonas titanicae was isolated from a culture of an alga strain, Spirulina platensis, and identified through 16S rRNA gene analysis. Corrosion behavior of 304L stainless steel (SS) coupons in the presence and absence of H. titanicae was characterized by using electrochemical measurements and surface analysis. The results showed that H. titanicae significantly accelerated the corrosion rate and decreased the pitting potential of 304L SS in the biotic medium. After removal of the corrosion products and biofilms, severe pitting corrosion caused by H. titanicae was observed. The largest pit depth after 14 d reached 6.6 μm, which was 5.5 times higher than that of the sterile control (1.2 μm). This is the first report revealing that an alga associated bacterium can induce microbiologically influenced corrosion (MIC), and a further concern is raised that whether algae play a role in the MIC process.
Keywords:
Recently, the shipping industries are continually facing the corrosion challenges caused by algae and microorganisms in the marine environment, which cause a substantial amount of economic damage [1]. The corrosion behaviors of algae have been well described in many reports [2]. It is well known that algae and bacteria coexist, and algae associated bacteria make a significant contribution to the growth and survival of algae [3]. However, little is known about the corrosion effect of the bacteria that coexist in symbiotic relationships with algae.
In the marine environment, biofilm formation is initiated by pioneer bacteria which colonize almost all surfaces and provide suitable and attractive conditions for the adhesion of algae and different micro- and macroorganisms [[4],[5]]. In these biofilms, algae produce oxygen and organic substrates, which can be used by heterotrophic bacteria in close proximity. In return, they can nutritionally benefit from compounds produced by these bacteria, such as vitamins and carbon dioxide [6]. These interactions are the primary cause of biofouling that leads to serious damage to marine materials [7]. Indeed, biofilm development and extension across the material surfaces can lead to changes in the physio-chemical conditions at metal/biofilm interface, which generally result in degradation of these materials in the form of localized corrosion [8]. This phenomenon is known as microbiologically influenced corrosion (MIC), and it is responsible for the severe economic loss that costs more than US$ 100 million per year to the oil and gas industry [9]. Based on the research of algal corrosion, it is crucial to further explore the corrosion behavior of symbiotic bacteria for marine corrosion prevention and control.
The present study focuses on an alga associated bacterium, Halomonas titanicae, which was first isolated and identified from rusticle samples formed by a consortium of microorganisms that was collected from the wreck site of RMS Titanic [10]. In this work, H. titanicae was isolated from a culture of Spirulina platensis, an alga collected from the Yellow Sea (China). Up to date, there is no report about the corrosion effect of H. titanicae and the role of this heterotrophic bacterium in the symbiotic relationship with an alga.
304L stainless steel (SS) is one of the most widely used materials in various applications because of its good corrosion resistance which is attributed to the chromium oxide film formed on its surface. However, it was reported that localized corrosion in the form of pits was observed underneath the biofilms on the 304L SS surfaces. Zhang et al. [11] reported that Desulfovibrio vulgaris increased the pit width and depth of 304 SS in the presence of electron mediators. In this work, we evaluated the corrosion behavior of 304L SS in the presence of H. titanicae biofilms in a simulated seawater environment by electrochemical and surface analysis techniques.
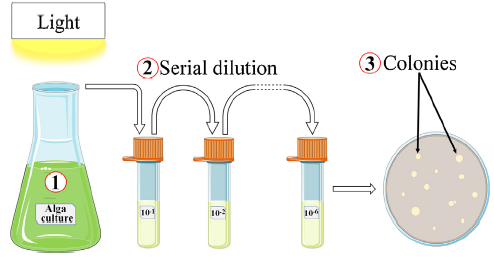
The alga strain, S. platensis, was obtained from the Marine Biological Culture Collection Center, Institute of Oceanology, Chinese Academy of Sciences (Qingdao, China). S. platensis was grown aerobically in F/2 medium [12]. In order to isolate the alga associated bacteria, a volume of 1 mL of the alga culture was used to inoculate into 200 mL of Luria-Bertani (LB) medium and further incubated under shaking for 24 h at 30 °C as shown in Fig. 1. Next, a series of decimal dilution were made by using PBS solution as a diluent, and a final volume of 100 μL was plated on LB agar plates. The appeared colonies were further purified by streaking on new LB agar plates.
Fig. 1. Schematic illustration of isolation and identification of the alga associated bacteria from S. platensis culture.
The bacterial isolate was identified by 16S rRNA sequencing [13]. The 16S rRNA sequence of the isolate was amplified by Sangon Biotech Co., Ltd. (Shanghai, China), and the amplified product was purified and sequenced by the same company. The obtained sequence was compared to the reported 16S rRNA sequence in the database of GenBank with the help of the BLASTn program. After that, the purified isolate was cultivated aerobically in 2216E medium (Qingdao Hope Bio-technology Co., China), which was based on artificial seawater (NaCl 19.45 g/L, MgCl2 5.98 g/L Na2HPO4 0.008 g/L, Na2SO4 3.24 g/L, CaCl2 1.8 g/L, KCl 0.55 g/L, Na2CO3 0.16 g/L, KBr 0.08 g/L, SrCl2 0.034 g/L, H3BO3 0.022 g/L, Na2SiO3 0.004 g/L, NaF 0.0024 g/L, NH4NO3 0.0016 g/L, peptone 5.0 g/L, ferric citrate 0.1 g/L and yeast extract 1.0 g/L). The MIC tests were conducted at 37 °C and the initial cell concentration was adjusted to 106 CFU mL-1 by using a UV-vis spectrophotometer. In addition, all the tests were conducted in two medium systems, the sterilized 2216E medium was called as sterile medium, while the medium inoculated with bacteria was named as biotic medium. There were three parallel samples in total for each test, and all the experiments were repeated thrice at least.
Square-shaped 304L SS coupons with sides of 10 mm and thickness of 1 mm were used. The chemical element composition of 304L SS specimens was obtained from the Department of Materials Analysis and Testing, Institute of Metal Research (Shenyang, China). 304L SS is composed of 18.95 wt% Cr, 0.02 wt% C, 1.11 wt% Si, 7.93 wt% Ni, 1.36 wt% Mn, and the balance of Fe. The specimens before MIC tests were mechanically abraded by silicon carbide papers from 400, 600, 800, 1000 to 1200 grits. For the electrochemical tests, the working electrodes were made according to the method previously described by Lekbach et al [14]. Before each experiment, the coupons were sterilized by immersing in 75% ethanol and exposing to UV light for 1 h.
The coupons immersed in the sterile and biotic media were taken out after 14 d of incubation. Biofilms (Live and dead cells) were stained and dehydrated by following the procedure previously described by Dong et al [15]. Then, the coupon surfaces were observed using confocal laser scanning microscopy (CLSM, C2 Plus, Nikon, Tokyo, Japan) and scanning electron microscopy (SEM, Ultra-Plus, Zeiss, Germany), respectively. To examine the pits formed on the coupon surfaces after the immersion test, the corrosion products and the biofilm were removed according to the GB/T4334.4-2000 standard (Chinese National Standards). The coupons were washed with distilled water and pure ethanol for 15 min in the ultrasonic bath and then immersed in the solution containing 10 mL HNO3 (analytically pure), 2 mL HF (analytically pure) and 100 mL distilled water for 5 min. Finally, the coupons were cleaned with distilled water again. After that, the coupons were cleaned with pure ethanol for three times, each time for 20 min in the ultrasonic bath (DSA100-SK2, Desen, China) and then dried in warm air. Next, the pit morphologies and depths were observed by CLSM (LSM 710, Zeiss, Germany). Moreover, the corrosion products on the coupon surfaces after incubation in the sterile and biotic media were characterized by X-ray photoelectron spectroscopy (XPS) (ESCALAB 250, Thermo Fisher, MA, USA).
The purified isolate was inoculated into electrochemical cells containing 300 mL of 2216E medium. Then, the electrochemical measurements were conducted every day during the 14 d period with the help of a potentiostat (Reference 600™, Gamry Instruments Inc., Warminster, PA, USA). Saturated calomel electrode (SCE) and platinum plate served as reference and counter electrodes, respectively. Open circuit potential (OCP) was first recorded as a thermodynamic parameter to explore the corrosion tendency. Linear polarization resistance (LPR) measurements were performed at a scan rate of 0.125 mV s-1 over the range of -10 to 10 mV versus the EOCP, with a sampling period of 1 s. Electrochemical impedance spectroscopy (EIS) was conducted at a steady state EOCP with an AC sine wave of amplitude 5 mV at frequencies ranging from 100,000 to 0.01 Hz. Furthermore, ZSimpWin software (V. 3.30d, EChem Software, USA) was used for EIS data analysis. Finally, potentiodynamic polarization curves were obtained at a scan rate of 0.5 mV s-1 and the potential range was from -500 to 1500 mV with respect to EOCP.
The bacterial isolate was identified by amplifying and sequencing the 16S rRNA gene. Molecular characterization revealed that the bacterial isolate (1478 bp) was phylogenetically related to members of Halomonas genus which belongs to the family of Halomonadaceae. The 16S rRNA sequence analysis of the isolate showed 99.9% identity with H. titanicae BH1. This bacterium is a rod-shaped, Gram-negative and heterotrophic. Moreover, it can grow aerobically in concentrations of NaCl ranging from 0.5% to 25% [10].
Fig. 2(a) shows the variation of EOCP values in the biotic and sterile media for 14 d. The OCP in the presence of H. titancae decreased drastically from -174 to -610 mV (vs. SCE), while OCP in the sterile medium dropped from -57 to -107 mV (vs. SCE). A similar trend was observed for 304L SS coupons in the presence of the corrosive bacterium, Pseudomonas aeruginosa, as reported by Lekbach et al [16]. They found that EOCP values decreased from -181 to -599 mV (vs. SCE) after 1 d of incubation. The EOCP of 304L SS coupon in the biotic medium shifted more negatively compared with that in the sterile medium, indicating a stronger tendency toward corrosion in the presence of H. titancae.
Fig. 2. (a) EOCP and (b) 1/Rp vs. exposure time for 304L SS coupons in the sterile and biotic media for 14 d, (c) comparisons of impedance values for 304L SS coupons in the sterile and biotic media after exposure for 1, 7 and 14 d and (d) comparisons of potentiodynamic polarization curves for 304L SS coupons after 7 and 14 d exposed to the sterile and biotic media, respectively.
The corrosion rate and tendency were investigated using electrochemical techniques. The corrosion rate was represented by the reciprocal of the polarization resistance (1/Rp) qualitatively, which was obtained from the LPR measurements [17]. Fig. 2(b) shows that corrosion rate increased rapidly after 24 h of incubation and reached the highest value of 22.2 μS cm-2 on the 3rd d. Then, it remained relatively stable until 21.4 μS cm-2 on the 9th d. Due to the death of the sessile bacteria, the corrosion rate decreased from the 10th d and reached the minimum value of 10.5 μS cm-2 on the 12th d, while the values of corrosion rate were quite stable at approximately 1.1 μS cm-2 in the sterile medium during 14 d. The results manifested that the corrosion rate in the presence of H. titancae was consistently higher than that in the absence of bacteria, and the average corrosion rate in the biotic medium (18.2 ± 1.3 μS cm-2) was 16.4 times larger than that in the sterile medium (1.1 ± 0.4 μS cm-2), wherein biofilms played a vital role to cause MIC.
Fig. 2(c) shows the impedance loops for 304L SS coupons in sterile and biotic media after exposure for 1, 7 and 14 d. In the Nyquist plots, the diameters of impedance loops markedly decreased with time from the 1st d to the 7th d in the biotic medium, implying the increase in the corrosion rate of 304L SS coupons. Moreover, the impedance loops increased with exposure time after 7 d suggesting the corrosion rate decreased, which was in accordance with results mentioned above [18]. On the contrary, the impedance loops in the sterile medium increased firstly and then decreased on the 14th d, which indicated that the corrosion rate was slow but increased with exposure time. In the presence of biofilm, the diagrams present two capacitive loops: the first loop of impedance plots was featured with a relatively small-flattened loop, which indicated that 304L SS electrode surface might become weak and porous. The high-frequency part showed the intact part of the formed film, while the low-frequency points indicated Faradaic process occurred on the surface of the 304L SS electrode through defects and pores in the adsorbed layer. In this case, the impedance plots confirmed that in the biotic medium, the corrosion mechanism contained two absorbed intermediates, the inherent passive film and the additional biofilm in this work [19]. The fitting parameters of corresponding equivalent electrical circuits are listed in Table 1, wherein Rs(QdlRct) and Rs(Qb(Rb(QdlRct))) were used to fit the EIS data of the 304L SS coupons in the sterile and biotic media, respectively. Rs represents the resistance of the solution. Qb and Rb are the capacitance and resistance of biofilm and/or corrosion products, respectively. Rct represents the charge transfer resistance, and Qdl refers to the capacitance of the electrical double layer (EDL). The Rct values in the biotic medium were two orders of magnitude lower than those in the sterile medium, indicating that the corrosion rate was accelerated in the presence of H. titanicae, which was in agreement with the LPR results shown in Fig. 2(b).
Table 1 Parameters extracted from EIS measurements for 304L SS in the sterile and biotic media during the incubation of 14 d.
| Day | Rs (Ω cm2) | Qf (×10-6 Ω-1 cm-2 Sn) | nf | Rb (Ω cm2) | Qdl (×10-6 Ω-1 cm-2 Sn) | ndl | Rct (×102 kΩ cm2) | X2 (×10-3) |
|---|---|---|---|---|---|---|---|---|
| In the sterile medium | ||||||||
| 1 | 13.9 | 21.9 | 0.91 | - | - | - | 35.9 | 1.0 |
| 4 | 14.3 | 20.9 | 0.91 | - | - | - | 61.7 | 1.6 |
| 7 | 14.4 | 20.4 | 0.91 | - | - | - | 46.9 | 1.5 |
| 10 | 14.0 | 20.9 | 0.91 | - | - | - | 38.3 | 1.1 |
| 14 | 14.8 | 24.4 | 0.90 | - | - | - | 34.0 | 1.0 |
| In the biotic medium | ||||||||
| 1 | 10.5 | 20.0 | 0.92 | 15.4 | 32.1 | 0.84 | 0.70 | 0.99 |
| 4 | 11.1 | 35.5 | 0.85 | 58.0 | 12.1 | 0.90 | 0.49 | 1.9 |
| 7 | 10.1 | 42.9 | 0.83 | 135 | 7.4 | 0.95 | 0.51 | 2.4 |
| 10 | 10.0 | 41.7 | 0.83 | 162 | 7.9 | 0.95 | 0.57 | 2.1 |
| 14 | 10.0 | 38.4 | 0.82 | 291 | 9.7 | 0.92 | 0.82 | 1.3 |
Kinetic corrosion parameters were obtained from polarization curves after 7 and 14 d of incubation in both sterile and biotic media. The oxygen deficiency caused by the oxygen consumption of H. titanicae biofilms may enhance the electron-transfer process from the anodic sites located underneath the H. titanicae biofilms to the cathodic sites where oxygen was reduced, resulting in the negative shift of EOCP (Fig. 2(a)) and polarization curves [20]. As shown in Table 2, the corrosion potential (Ecorr) decreased from -0.20 V (vs. SCE) on the 7th d to -0.26 V (vs. SCE) on the 14th d in the sterile medium. Whereas, the Ecorr values in biotic medium reached -0.79 V (vs. SCE) after 14 d. This result showed that the corrosion tendency was considerably higher in the presence of H. titanicae and increased with exposure time. The corrosion current icorr in the biotic medium increased from 2.4 μA cm-2 on the 7th d to 5.8 μA cm-2 on the 14th d. In comparison, the icorr value in the sterile medium decreased and reached 0.3 μA cm-2 after 14 d. This result confirmed that the corrosion rate was accelerated in the presence of H. titanicae, which corroborated the LPR and EIS data. It has been found that microorganisms mainly cause pitting corrosion [21]. The pitting potential (Epit) value decreased drastically to 0.26 V (vs. SCE) in the biotic medium, while the Epit in the sterile medium was 0.70 V (vs. SCE) after 14 d, demonstrating that H. titanicae significantly affected the pitting potential of 304L SS with increased immersion time. The decrease of pitting potential in the presence of H. titanicae also indicated that 304L SS was particularly prone to MIC pitting caused by the H. titanicae biofilm.
Table 2 Corrosion parameters from the polarization curves for 304L SS in the sterile and biotic media for 7 and 14 d.
| Medium | Duration (d) | icorr (μA cm-2) | Ecorr (V) vs. SCE | Epit (V) vs. SCE |
|---|---|---|---|---|
| Sterile medium | 7 | 0.5 | -0.20 | 0.45 |
| 14 | 0.3 | -0.26 | 0.70 | |
| Biotic medium | 7 | 2.4 | -0.69 | 0.41 |
| 14 | 5.8 | -0.79 | 0.26 |
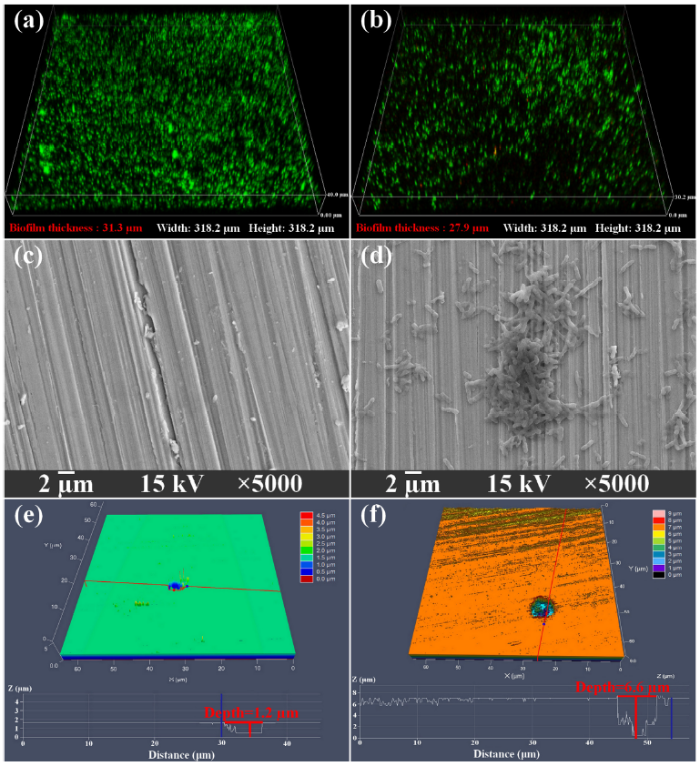
It is widely accepted that biofilms are responsible for MIC [22]. Biofilms grown on the 304L SS coupon surfaces were observed after incubation time (Figs. 3(a), (b) and (d)). The maximum biofilm thickness after 7 d was 31.3 μm with an average thickness of 29.8 ± 1.4 μm and then it decreased to 27.9 μm with an average thickness of 26.6 ± 1.3 μm after 14 d. This result was consistent with electrochemical analyses and confirmed that the corrosion rate acceleration of 304L SS was due to the formation of the biofilms. After removal of the biofilms and corrosion products on the coupon surfaces, MIC pits were found as shown in Figs. 3(e) and (f). The largest pit depth was 6.6 μm with a width of 6.9 μm in the biotic medium, while the largest pit depth was only 1.2 μm with a width of 6.3 μm in the sterile medium. Meanwhile, the average pit depths were 4.6 ± 1.7 μm and 1.1 ± 0.1 μm in the biotic and sterile media, respectively. This confirmed that 304L SS coupons were vulnerable to MIC in the presence of H. titanicae, and the corrosivity of H. titanicae was close to the corrosive P. aeruginosa as reported by Jia et al. [23]. They found that the largest pit depth on 304 SS surface after 14 d of incubation in anaerobic conditions was 7.4 μm. In the worst-case scenario (the MIC rate is assumed to be a constant), the H. titanicae can trigger a corrosion rate of 1.7 mm/a, which is fatal to the 304L SS.
Fig. 3. CLSM 3-D images of H. titanicae biofilm on the 304L SS coupon surfaces after (a) 7 d, (b) 14 d; SEM images of the 304L SS coupon surfaces after exposure to (c) sterile medium and (d) biotic medium for 14 d; the largest pit depth on 304L SS coupon surfaces after 14 d of exposure to (e) sterile medium and (f) biotic medium.
Corrosion products on the 304L SS coupon surfaces were analyzed after 14 d (Fig. 4). XPS spectra revealed that there was no significant difference in corrosion products on the coupon surface in the abiotic and biotic media. The corrosion products were mainly Fe (707.0 eV), Fe2O3 (710.7 eV) and FeOOH (724.3 eV). The latter was unstable and continued to be decomposed to Fe2O3 [24]. In addition, the high-resolution spectra of Cr 2p confirmed two peak components for the abiotic control with Cr2O3 (576.7, 586.0 eV) and Cr(OH)3 (577.0 eV), while only Cr2O3 (586.2 eV) was detected in the presence of H. titanicae. Li et al. [25] reported the similar phenomenon that Cr2O3 was found under the aerobic biofilm. The process could be expressed in equations as follows [[26],[27]]:
Anodic oxidation: Fe → Fe2+ + 2e- (1)
Cathodic reduction: 1/2O2 + H2O + 2e- → 2OH- (2)
Ionic reaction: Fe2+ + 2OH- → Fe(OH)2 (3)
Oxidizing reaction: 2Fe(OH)2 + 1/2O2 → 2FeOOH + H2O (4)
Decomposition reaction: 2FeOOH → Fe2O3 + H2O (5)
Fig. 4. XPS spectra of Fe 2p, O 1s and Cr 2p for 304L SS coupons after 14 d incubation in the sterile medium (a, c, e) and biotic medium (b, d, f).
The pH values in abiotic control changed slightly and reached 7.0 after 14 d, while the pH in the presence of H. titanicae even increased to 8.1, indicating that acid attack caused by the metabolism of H. titanicae was impossible. The H. titanicae aerobic biofilms formed on the coupon surfaces may change the concentration of oxygen and ions, even pH (hydrolysis of excess iron ions) underneath the biofilms, and thus caused the severe MIC of 304L SS. Besides, aerobic biofilms were proven to induce the MIC by accelerating the electron transfer rate between the metal matrix and bacterial cells via production of electron meditators, as Huang et al. reported in their study [28]. This mechanism is called extracellular electron transfer (EET)-MIC. The anodic reaction was accelerated by the biofilms as shown in Fig. 2(d), which could attribute to the enhancement of EET efficiency between the cell wall of H. titanicae and SS matrix. The electron uptake can be achieved by either mediated electron transfer (MET, electron shuttles) or direct electron transfer (DET, c-type outer-membrane cytochromes and pili). The mechanism and EET pathway of H. titanicae should be further investigated.
In this work, the electrochemical and surface analysis results showed that H. titanicae induced severe MIC attack on the 304L SS. The corrosion rate in the biotic medium (18.2 ± 1.3 μS cm-2) was 16.4 times higher than that in the sterile medium (1.1 ± 0.4 μS cm-2), and the largest pit depth after 14 d reached 6.6 μm, which was 5.5 times deeper than that of the sterile control. The formation of aerobic biofilms on the coupon surfaces may result in EET-MIC. In addition, due to the mutually beneficial symbiosis of algae and its associated bacteria, it is difficult to separate them. The corrosivity of algae against metal materials applied in the deep-sea cage, geopolymer concrete and other marine infrastructures has been reported [29]. They were persistently regarded as the sole contributor to the MIC process. Therefore, we raise a concern that whether the reported corrosion caused by the algae was indeed due to algae alone, or algae may even not be a player in the corrosion process. Further study and more evidence are needed to elucidate the roles of algae and its associated bacteria in the biocorrosion process.
This work was supported financially by the National Natural Science Foundation of China (Nos. U1660118 and 51871050), the National Environmental Corrosion Platform (NECP) of China, and the Fundamental Research Funds for the Central Universities of Ministry of Education of China (No. N180205021).

 WeChat
WeChat
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}